Area professionisti | Le schede coltura | La fertilizzazione | Fisiologia e nutrizione
Area professionisti | Le schede coltura | La fertilizzazione | Fisiologia e nutrizione
La nutrizione del ciclamino è condizionata innanzi tutto da fattori climatici e dalla disponibilità di ossigeno e di acqua per le radici. Questi parametri portano ad una selettività a livello radicale e modificano la dinamica degli elementi nutritivi nella pianta.
L’analisi del corretto o incorretto sviluppo del vegetale si basa su una buona conoscenza del suo ciclo vegetativo.
Ai fini di una precisa analisi, però, occorre prendere determinate chiavi e aprire le porte nell'ordine giusto.
E', infatti, necessario capire a fondo la funzione di ogni chiave, ossia di ogni fattore di produttività vegetale, e conoscere il suo ruolo all'interno del ciclo vegetativo.
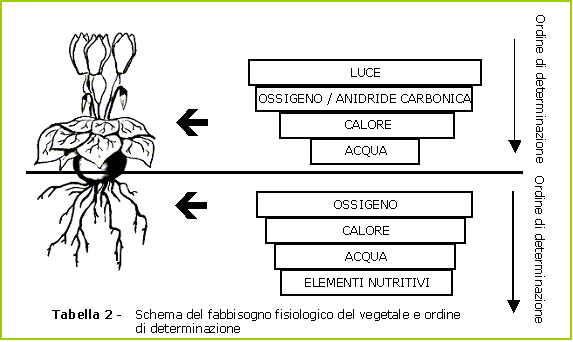
Il mondo delle piante è strutturato in maniera tale che alcuni fattori sono più rapidamente «limitanti» di altri. In particolare, è inutile cercare di fissare un equilibrio nutritivo finché i fattori primi, ossia luce, aria, calore e acqua, continuano a svolgere un ruolo limitante.

Esempio:
La radice, respirando, esercita un'assimilazione attiva degli elementi nutritivi.
In caso d'insufficienza respiratoria legata ad una pressione dell'ossigeno troppo bassa, i meccanismi di assimilazione diventano passivi. La radice non è più in grado di lottare attivamente contro l'assimilazione di «elementi tossici».
La radice s'intossica o dà luogo alla formazione di «barriere» di protezione, dette «bruciature», impedendo la penetrazione eccessiva di elementi nutritivi che, poiché la radice non è più in grado di respirare, non sarebbero metabolizzabili.
La luce è il primo fattore indispensabile alla vita vegetale. Essa, infatti, fornisce l’energia necessaria al funzionamento del motore vegetale.
L’ossigeno e l'anidride carbonica costituiscono il secondo fattore di produttività dopo la luce.
Essi rappresentano, infatti, la base stessa della sostanza vegetale, composta per la maggior parte da:
|
: |
C, proveniente da CO2 (anidride carbonica) |
|
: |
O, proveniente da O2 (ossigeno) |
|
: |
H, proveniente da H2O (acqua) |
Contrariamente a ciò che si pensa, l'ossigeno, a livello delle radici, è un fattore limitante ancor prima dell'acqua. Il ciclo vegetale del ciclamino si basa infatti su una serie di reazioni a dominante ossidante.
La radice, in presenza di una buona pressione di ossigeno, sviluppa dei meccanismi attivi di assimilazione. Essa, quindi, è in grado di selezionare gli elementi nutritivi di cui ha bisogno e di trasferirli in maniera attiva verso gli organi di destinazione.
Se il fabbisogno di ossigeno delle radici non viene soddisfatto, la radice sviluppa temporaneamente reazioni «passive» a dominante riducente.
Queste reazioni passive sono reazioni di «compensazione» messe in atto nell'attesa che l'ossigeno torni ad essere il fattore più limitante.
L’inconveniente di queste reazioni «passive» risiede nel fatto che il ciclamino s’intossica. La pianta, infatti, non è più in grado di esercitare un controllo «attivo» sulla propria nutrizione.
In questa fase sono assimilati, in particolare, l'azoto ammoniacale, l'alluminio, il manganese…
Queste forme «ridotte» penetrano passivamente nella radice. Quest'ultima, non potendo svolgere una normale attività ossidante, accumula tali forme ridotte che, naturalmente, la intossicano. Appaiono allora malformazioni sulle foglie e punture sui fiori (eccesso di manganese). Di lì a poco, il ciclamino mette in atto dei freni "anti-veleno", ossia sviluppa sulle radici delle forme sugherose che fungono da barriera di difesa.
La perdita delle radici, a questo punto, è imminente.
Questi fenomeni passivi cessano quando la pressione dell'ossigeno torna a valori normali. Gli elementi tossici assimilati vengono allora ossidati e «neutralizzati».
Tuttavia, si tratta soprattutto di una nozione di «flusso» di ossigeno più che di una nozione di quantità.
Tutto dipende dalla velocità alla quale è soddisfatta la pressione «normale» dell'ossigeno.
Questa nozione di velocità di rinnovamento dell'ossigeno sarà uno degli elementi decisivi per la scelta del substrato.
Si noti, inoltre, che qualsiasi attività «ossidante» nella radice genera un ambiente riducente all'interno del terreno colturale. Ciò significa che, quando la radice respira e consuma parte dell'ossigeno sciolto in soluzione, rilascia anche anidride carbonica (CO2).Da qui la nozione di flusso e di circolazione dei gas, aspetto che assume un'importanza primordiale a livello della radice.
L’incremento della pressione parziale dell'anidride carbonica attorno alle radici (residuo della respirazione) avviene a danno della pressione parziale dell'ossigeno.
L'unico modo per mantenere una pressione dell'ossigeno sufficiente consiste nell'espellere l'anidride carbonica.
L’azione di «espulsione» è svolta essenzialmente dalla circolazione dell'acqua.
Di conseguenza, è inutile dare da bere ad una pianta che ha freddo e ancor più inutile e pericoloso è nutrirla.
Si noti che il binomio annaffiatura/substrato, condizionando innanzi tutto la pressione dell'ossigeno disponibile per le radici, condiziona anche il «calore» fornito a livello delle radici.
Esempio:
Caso di rose riprodotte per talea, coltivate su substrati a base di torbe fini, abasso contenuto di fibre grossolane. Annaffiatura con acqua non riscaldata e fresca - stagione invernale.
Ogni annaffiatura superiore alla conducibilità di 1,5 mS/cm provoca bruciature sulle radici.
Il fattore limitante, in questo caso, è il calore fornito alle radici.
Il substrato presenta un calore specifico insufficiente. «L’aria» calda non circola a sufficienza, in quanto il substrato rimane inzuppato d'acqua troppo a lungo.
La radice, allora, abbassa la propria soglia di tolleranza alla conducibilità.
Una volta soddisfatti i primi tre fattori, il ciclamino può finalmente nutrirsi!
La caratteristica principale della nutrizione idrica risiede nel fatto che si tratta soprattutto di una nutrizione di transito.
L'acqua, infatti, penetrando attraverso le radici, è rilasciata nell'atmosfera tramite la traspirazione degli organi aerei, in quantità di gran lunga superiore a quella che rimane nel ciclamino a titolo permanente.
La nutrizione minerale riguarda tutti gli elementi nutritivi diversi da C, O, H.
Essa è caratterizzata da un'incorporazione più o meno completa di questi elementi nutritivi nella sostanza vegetale del ciclamino.
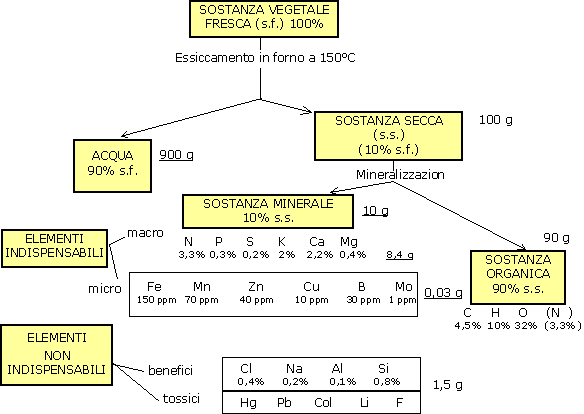
Gli elementi minerali sono generalmente assorbiti sotto forma di ioni, prodotti dalla decomposizione nell'acqua degli elettroliti e dei sali minerali (concimi).
Esistono però anche altre molecole, diverse dagli ioni, che possono essere assimilate dalla pianta:
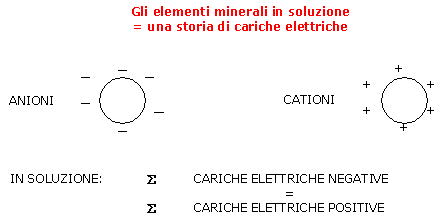
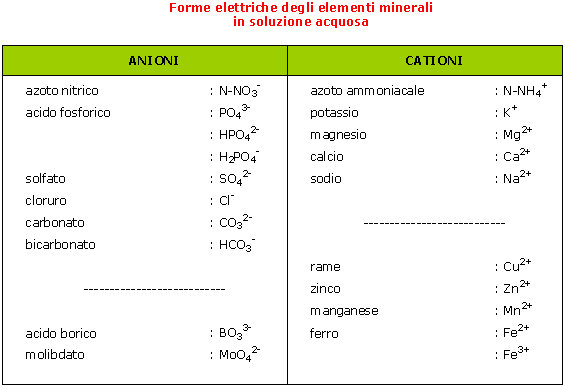
In soluzione, la somma delle cariche elettriche negative è sempre uguale alla somma delle cariche elettriche positive.
Gli organi di assorbimento sono formati dal complesso delle radici, talvolta dalla foglia ed eccezionalmente dal frutto o dal gambo.
Innanzi tutto va ricordato che la nutrizione per via fogliare non può costituire in nessun caso una via di nutrizione permanente. Essa deve essere considerata solo nell'ambito delle polverizzazioni preventive (in particolare prima di alcuni stadi specifici) o come «rimedio». In tal caso, infatti, essa interviene al momento giusto per correggere uno squilibrio nutritivo, ma non può comunque essere in grado di fornire alla pianta tutti gli elementi di cui essa ha bisogno.
Al contrario, la principale via di assorbimento dell'acqua e degli elementi minerali è rappresentata dai peli assorbenti delle giovani radici.
L'assorbimento, infatti, è condizionato essenzialmente dall'esistenza di barriere al passaggio di tali sostanze nel ciclamino.
Il pelo assorbente possiede una membrana molto sottile che consente una facile diffusione degli elementi.
MA ATTENZIONE! C'è pelo assorbente e pelo assorbente!
Talvolta il ciclamino è costretto a produrre lunghe radici immature che tendono ad arrotolarsi o a svilupparsi in maniera irregolare. Queste radici cercano il contatto con il substrato che non trovano, e la pianta continua allora a sviluppare radici fin quando non si crea questo contatto.
(E' il caso dei terreni colturali a porosità molto grossolana).
Queste radici sono immature, non suberificate, ma non costituiscono in nessun caso un «lungo» pelo assorbente.
In una radice attiva, la lunghezza efficace in termini di «capacità di assorbimento» è di soli 2 o 3 centimetri.
Attenzione anche alle radici che presentano assise suberificate (aspetto sugheroso, un po' spesso). Tali assise costituiscono una barriera per il passaggio dell'assorbimento di base e quindi la «capacità di assorbimento» di questo tipo di radici è ridotta.
Il ciclamino, come tutte le specie bulbose, è classificato come pianta "esigente" ai fini della determinazione della fertilizzazione.
Le piante bulbose presentano, infatti, un potenziale di estrazione degli elementi nutritivi tra i più bassi.
Inoltre, i bulbi e il loro sistema radicale presentano due caratteristiche antagoniste: sono esigenti in termini di ossigeno ma non sopportano i terreni colturali a porosità troppo grossolana che limitano il contatto radice/bulbo/substrato e accrescono quindi la difficoltà per la radice di estrarre gli elementi nutritivi, come il fosforo e gli oligoelementi.
I casi di carenze osservati sul ciclamino sono frequenti: carenze di boro, di manganese e di ferro. Essi sono legati, in particolare, ad uno scarso contatto tra la radice e il substrato.
Questo debole potere di estrazione del sistema radicale del ciclamino è alla base della sua classificazione come specie esigente in oligoelementi.
Esempio:
La nutrizione fosforica è da rapportare esclusivamente alla capacità del ciclamino nel rinnovare i peli assorbenti.
Alcuni studi hanno dimostrato che solo i peli assorbenti sono in grado di assimilare il fosforo presente nel terreno colturale, soprattutto nelle fasi più giovani della rizogenesi. Di conseguenza, è solo la frazione di fosforo solubile nell'acqua ad essere assimilata.
L’aumento di uno degli elementi in prossimità della radice o del substrato provoca una fissazione o un ulteriore assorbimento di detto elemento a danno degli altri, a condizione che le loro concentrazioni non siano diminuite!
L’adsorbimento, fenomeno tramite il quale i solidi o le soluzioni trattengono sulla loro superficie molecole o ioni in fase gassosa o liquida, non deve essere confuso con l'assorbimento, vale a dire la proprietà dei solidi o dei liquidi di trattenere determinati gas o liquidi in tutto il loro volume.
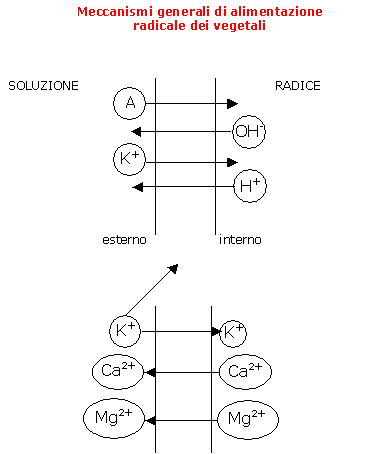
Le membrane cellulari sono ricche, in particolare, di poli elettronegativi. Esse possiedono, quindi, una C.S.C. radicale, ossia una Capacità di Scambio Cationico, in pratica la capacità di fissare i cationi della soluzione.
I cationi del terreno colturale sono adsorbiti su questa C.S.C. radicale in cambio di un catione interno alla cellula che, a sua volta, è rilasciato nel terreno colturale (cfr. schema).
E' il caso dei seguenti cationi: K+ (potassio), Ca2+ (calcio), Mg2+ (magnesio), Na+ (sodio), NH4+ (ammonio).
Nel caso degli oligoelementi, il meccanismo è diverso.
Gli oligoelementi sono presenti raramente in forma ionica nella soluzione. Il più delle volte essi sono «chelati» sulla sostanza organica tramite piccoli chelati organici.
In questo caso, è evidente che potranno essere assimilati dalla radice solo se il pelo assorbente è a contatto fisico con il supporto colturale (terreno, substrato, ...). Questo contatto tra il chelato organico e la C.S.C. (poli negativi) del pelo assorbente consente di adsorbire, ovvero di fissare, l'oligoelemento (Cu2+, Mn2+, Zn2+, Fe2+).
L’assorbimento e l'adsorbimento sono meccanismi passivi, per i quali, cioè, la pianta non consuma energia.
Per continuare l'adsorbimento oltre lo spazio libero delle radici, la pianta mette in atto dei meccanismi attivi: il pompaggio metabolico.
Questa fase consente agli elementi nutritivi di penetrare in senso opposto al loro gradiente di concentrazione.
Essa, inoltre, consuma grandi quantità di energia, liberata dal processo di respirazione.
Col termine respirazione intendiamo il fabbisogno di ossigeno a livello aereo e a livello radicale!
Ecco perché un ciclamino «asfissiato» non può alimentarsi.
Il ciclamino seleziona gli elementi minerali. Questa selettività radicale avviene all'interno della pianta, secondo due principi :

Si noti, in questo esempio, che il potassio e il sodio hanno pressoché le stesse dimensioni. Il sodio, quindi, può sostituirsi facilmente al potassio.
|
Ca2+ |
> |
Mg2+ |
> |
K+ |
> |
Na+ |
> |
H+ |
Unendo queste due leggi, si ottiene il seguente ordine reale di velocità di penetrazione delle membrane:
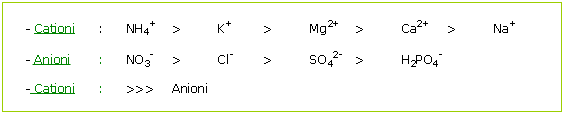
Si osservi che il potassio e l'azoto penetrano molto velocemente e con la stessa facilità nelle membrane, motivando così l'interesso verso l'uso del potassio per neutralizzare «l’azoto».
Non appena si manifesta una carenza di azoto, la radice seleziona l'anione che penetra più velocemente dopo l'azoto, ossia i cloruri.
Attenzione, quindi, alle carenze di azoto nitrico in terreni colturali ricchi di cloruri.
Il magnesio e il calcio sono cationi di grosse dimensioni che possono diffondersi male nella radice, soprattutto quando quest'ultima è soggetta a fenomeni di stress di altra natura.
I fosfati penetrano con molta difficoltà nelle membrane, per cui è necessario ottenere peli radicali assorbenti di una certa potenzialità per compensare la cattiva assimilabilità da parte della radice.
La composizione di ogni ciclamino si differenzia, inoltre, a seconda della natura del terreno colturale sul quale cresce, e l'intensità di assorbimento di ogni singolo elemento nutritivo dipenderà in particolar modo dalla concentrazione dei rispettivi ioni nel terreno colturale.
La presenza, seppur generalizzata, di un elemento nell'ambiente naturale non significa che questo elemento sia indispensabile alla vita del ciclamino. L'assorbimento, infatti, non è un fenomeno intelligente. Nonostante la presenza di alcuni meccanismi di regolazione, i ciclamini sanno rifiutare gli elementi particolarmente tossici, o che diventano tali in caso di eccesso, solo in determinate condizioni e fino ad un certo limite.
Si distinguono:

Si noti che la nozione di tossicità è «arbitraria» se estrapolata dal suo contesto agronomico.
La tossicità, naturalmente, è una nozione di eccesso, ma l'interpretazione dei sintomi visibili è sempre molto discutibile.
L’azoto è il principale fattore di crescita dei gambi e delle foglie.
Il fosforo «solubile» in acqua favorisce lo sviluppo dei peli assorbenti, ma non ha alcun effetto «generatore» di radici, nel senso proprio del termine.
E’ sufficiente che una piccola frazione di fosforo contenuta nel terreno colturale sia presente sotto forma di fosfati solubili in acqua (H2PO4) affinché il giovane pelo assorbente possa assimilarla.
Questo fosforo è successivamente incorporato nelle cellule sotto forma di ATP (molecola energetica).
Questa energia accumulata consente alle giovani radici appena nate di continuare a crescere.
Non bisogna quindi confondere la quantità di fosforo apportata e la forma del fosforo presente nel terreno colturale! Evidentemente, le giovani radici esportano pochissimo fosforo, ma quest’ultimo deve essere immediatamente disponibile (nozione di solubilità delle forme fosfatate).
Perché questa posizione di «catione maestro» che fa di questo catione il principale agente di controllo della qualità dei vegetali?
Il potassio è l'elemento prediletto dagli organi di assorbimento, grazie alla sua grande diffusibilità completata da una buona selettività.
Il potassio fornito viene assimilato, persino in altissime quantità, poiché non esiste quasi nessun freno alla sua assimilazione, il che può provocare una scarsa attivazione del coefficiente di respirazione e affaticare il ciclamino.
Il potassio, inoltre, svolge funzioni particolarmente estese.
I tassi di concentrazione più frequenti nel ciclamino sono analoghi a quelli dell'azoto e spesso anche più elevati.
La materia vivente, nel corso del proprio ciclo vitale, produce sempre una quantità predominante di prodotti con cariche negative (radicali acidi o a carattere acido prodotti dal metabolismo).
Spesso il potassio è l'unico catione in grado di penetrare nelle cellule per neutralizzare questi radicali acidi (negativi) man mano che si sviluppa il loro processo di sintesi.
Per questo motivo si dice che il potassio «favorisce» la fotosintesi e stimola la respirazione, attivando quindi la formazione di riserve.
Il potassio, essendo altamente solubile, è il principale catione a influire sulla pressione osmotica delle cellule.
Più il potassio si concentra nei vacuoli delle cellule, più la pressione osmotica aumenta, e più il ciclamino assorbe acqua attraverso la radice per «diluire» questa concentrazione di potassio nella cellula.
Nasce da qui il ruolo di stimolatore dell'assorbimento d'acqua del potassio.
Il miglioramento della ritenzione idrica da parte del vegetale favorisce la resistenza al gelo e ad alcune malattie.
Il potassio, però, evita anche lo spreco di acqua tramite altre funzioni.
La più importante è quella che lo vede coinvolto nel movimento degli stomi (piccoli pori di apertura a livello delle foglie) che possono aprirsi o richiudersi e che condizionano la traspirazione.
Il potassio, infatti, riduce il periodo durante il quale l'apertura degli stomi non è adeguata alle condizioni del terreno colturale.
Questo spiega il suo ruolo nella resistenza alla siccità del ciclamino nel periodo estivo.
Il potassio attiva moltissimi enzimi e favorisce la sintesi dei composti polimerizzati.
Questo spiega l'influenza del potassio sulla sintesi dei pigmenti responsabili della colorazione dei fiori.
Il potassio svolge un effetto positivo principalmente per il trasporto degli zuccheri.
Questo spiega il ruolo del potassio nella creazione di riserve nel bulbo del ciclamino.
Il ruolo della fertilizzazione consiste quindi nel fornire una pressione potassica sufficiente a livello radicale circa 15 giorni prima dell'effetto desiderato.
Il calcio svolge un ruolo plastico, in quanto è parte integrante delle pareti cellulari pectocellulosiche.
Esso svolge anche una funzione antitossica: neutralizza gli ioni acidi (solfati, carbonati...) nelle «sottostrutture».
Il calcio è assai poco mobile all'interno della pianta.
Esso contrasta l'influenza del potassio.
Il potassio, infatti, può intossicare la pianta favorendo un'eccessiva permeabilità cellulare e il calcio compensa questa influenza diminuendo la permeabilità.
Nasce da qui la nozione di antagonismo K / Ca. Si noti che questo antagonismo K / Ca dipende essenzialmente dal regime idrico applicato al ciclamino. L'assimilazione del potassio è favorita da annaffiature poco frequenti effettuate con grossi moduli, mentre annaffiature molto frequenti e l'uso di piccoli moduli favoriscono l'assimilazione del calcio e del magnesio.
La molecola di clorofilla contiene un atomo di magnesio.
Questo elemento favorisce una fioritura precoce.
Attenzione alle confusioni!
Bisogna distinguere bene tra ciò che ci insegna la fisiologia vegetale e ciò che è in grado di fornire il terreno colturale.
Tuttavia, il fatto che esista un antagonismo tra K e Ca o tra K e Mg non significa che, all'analisi del substrato o del terreno, un rapporto K/Ca debba essere interpretato negli stessi termini.
Infatti nei substrati, che nella maggior parte dei casi sono ricchi di calcio e di magnesio, l'assimilazione dei tre cationi (K, Ca e Mg) non è regolata da relazioni di antagonismo, bensì dai livelli d'idratazione e del coefficiente di respirazione.

Quando A aumenta nel terreno colturale e B rimane costante, l'assorbimento di A aumenta, ma l'assorbimento di B diminuisce.
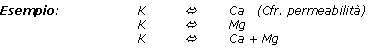
Il magnesio presenta un comportamento «intermedio» tra K e Ca. Nei substrati organici, la carenza di magnesio è provocata, il più delle volte, da un difetto della struttura o dell'aerazione del substrato che genera una difficoltà di assimilazione del magnesio. L'influenza della concentrazione di potassio nel terreno colturale è quasi nulla.

Quando aumenta la concentrazione dell'elemento A in soluzione e l'elemento B rimane costante, l'elemento A fa aumentare l'assimilazione dell'elemento B da parte della pianta.
Consideriamo qui di seguito due sinergie essenziali:
Questa sinergia svolge un ruolo importante in primavera nei terreni o nei substrati freschi. Il fatto di apportare in maniera congiunta fosfati e azoto favorisce, infatti, l'assimilazione del fosforo negli stadi più giovani della rizogenesi.
Questa sinergia ha luogo solo se è presente una frazione di fosfati solubile in acqua.
Nasce da qui l'interesse per apporti di concime a base di fosfato ammoniacale.
L'apporto di magnesio stimola l'assorbimento del fosforo.
In caso di situazione limitante, dovuta ad una perdita di radici, l'apporto di magnesio favorisce l'assimilazione del fosforo che spesso il sistema radicale stressato non è più in grado di assimilare efficacemente. Questo apporto previene la caduta delle foglie e favorisce l'attivazione delle cellule radicali.
La fertilizzazione deve stabilire un'offerta per consentire al ciclamino, assieme ai fattori che agiscono sulla nutrizione, di soddisfare il proprio fabbisogno interno.
Questo fabbisogno interno si manifesta attraverso l'esportazione di elementi nutritivi.
Tuttavia, non basta conoscere semplicemente la quantità delle esportazioni.
Bisogna aggiungere a questo dato dei coefficienti di maggiorazione che dipendono soprattutto dal terreno colturale.
2565, rue de Montourey
83600 Fréjus - France
Telefono internazionale : +33 (0)4 94 19 73 04
Centralino : + 33 (0)4 94 19 73 00
Fax : +33 (0)4 94 19 73 19